 |
Factoring Sequences
Van Warren Warren Design Vision 1996 |
Introduction
Proteins, the building blocks of life, can be represented as sequences of consecutive symbols. At a primitive level, the four nucleic acids represented by C, T, A and G code for a set of twenty primitives, the amino acids. The amino acidsare assembled into substructures with identifiable functional roles.
The goal of this note is to apply the concept of substructures - repeatable patterns and subunits - in an indirect way. Instead of attempting to deduce the function of substructures directly, this work seeks to catalog them by progressive abbreviation; To identify the frequency of occurrence of constituent fragments which are present in various proteins, enzymes, viruses, and genes. It is hoped that an indirect approach of this sort might lead to some insight about higher level function.
By identifying and counting repeating substructures it is hoped that some sort of a clue, some tiny insight might appear, which would yield value to an experienced worker, who would perhaps be able to deduce the role of that more complex assemblage. I got this idea from my work in finite element modeling where simple structures are repeatedly assembled into larger ones with more complex roles.
Questions
We take as a working example a ribosomal protein that plays a role in breast cancer, L-19. We begin by asking some statistical questions about L19:
1) What is the longest substring in L-19 that repeats? Call this substring S1.
2) How many instances of this substring are there? Call this count C1.
We continue by asking this question again in successively smaller increments, to wit:
3) What is the next longest substring in L-19 after S1? Call this substring S2.
4) How many instances of this substring are there? Call this count C2.
The telecommuting pioneer, Herb Younger of JPL once said, "When all you have is a hammer, everything looks like a nail." Applying his maxim we obtain:
5) What is the incidence of successively smaller substrings in L-19? We will number these substrings S3 - SN.
6) What frequency of incidence can we associate with substrings S3 - SN? Call this C3 - CN.
Answering these questions enables us to produce a table of all repeating substrings together with a frequency of each. Possessing this table would allow us to surmise what fraction of L-19 consists of things that repeat, and what fraction consists of things that are unique, thus this technique's name. On viewing this table, perhaps one might deduce some fact relating to substructure function, some kind of clue relating to purpose, simply by viewing the extent of repetition of various primitive substructure patterns at consectutively higher levels of organization;
Combinatorial Analysis
Before we go about answering the six questions above, we need to do some analysis. To "count the cost" prior to embarking on an experiment, we need to calculate how much computer memory and running time will be required to complete the operation for a given sequence. In computer science parlance this is called, "finding the space and time complexity" for the method.
First we observe that for a string S0 of arbitrary length C0 there are:
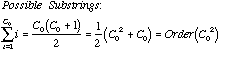
The term "Order" implies
the rate at which the number of substrings grows as a function of string
length.
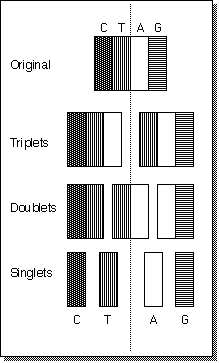
In the above case we see that the
growth is quadratic. This can be seen by looking at the diagram of a
trivial case, the nine unique substrings of CTAG - the title figure of
this article. Starting at the second row and working down we have 2 strings
+ 3 strings + 4 strings. We note that the total number of rows equals
the length of the original string which is consistent with the summation
expression given above.

Noting that we have cubic growth, we plot the expression and obtain the graph of space complexity:

Our analysis implies we must store all the substrings simultaneously. In fact we can generate them a row at a time, with a considerable savings of space. With a little cleverness we can make the storage requirement obey a linear growth law, meaning that there is a linear relationship between the length of the original input string and the storage required to generate the substrings. This is very desirable.
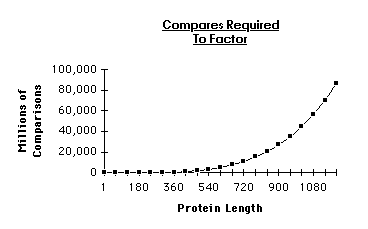
To obtain the execution time we must find the number of comparisons
required. To do this we compute the number of comparisons performed on
each row with the understanding that we compare all substrings on a given
row with each other to find repeating occurrences.
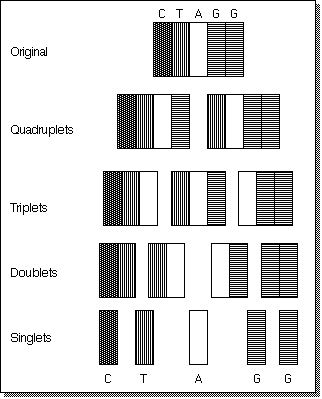
The figure above shows how a more complex string (chosen arbitrarily) would be broken into its possible substrings. The execution time complexity is the sum of the product of the comparisons per row and the length of the strings in that row:
![]()
More precisely we have:
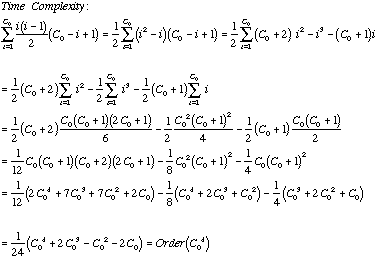
Plotting the time complexity we have:
facString Usage and Development
A 'C' program which implements this factoring algorithm was implemented. It reads the sequence of interest, factors it and prints the results.
Its usage is as follows:
facString sequenceFile
Sequence File Format:
sequence_length longest shortest
sequence
where:
sequence_length : an integer specifying the length of the sequence.
longest : the length of the longest repeating string to search for.
shortest: the length of the shortest repeating string to search for.
sequence: any sequence of letters representing nucleic or amino acids.
For an exhaustive search one would specify longest as (sequence_length - 1) and shortest as 1. It is often convenient to bracket the search by setting these to a narrower range of more specific values. This saves computer time. A consequence of the way facString is implemented is that if longest and shortest are both set to 1, facString just counts c, t's, a's and g's,
A couple of interesting facts emerged during the writing of facString; It is not necessary to make copies or subcopies of the original string in order to factor it; Further it is not necessary to declare a specific set of symbols as significant, that is what we're trying to find. For convenience the input is limited to upper and lowercase letters so that numbers can be used for annotation. Substrings are represented as a pair of integer coordinates indicating positional station along the string. The output is annotated to show:
1) the origin of the first occurrence of a repeating sequence and
2) the distance away that successive instances were found.
An advantage of knowing the distance is that it is then easy to determine at a glance whether a repeating unit occurs as part of the current unit (a negative distance), whether it abuts directly with the current string ( a zero distance), or whether it occurs further away ( a positive distance). This is illustrated below. These distances can then be plotted. As mentioned above the implementation makes it fast to search for repeaters of a specific length. It is handy (as in fun) to run facString and then read the output file into Microsoft Excel for subsequent analysis.
Output Format
The short string output format is:
string number_of_reps (first _location) first_distance sec_distance ...
This is depicted below.
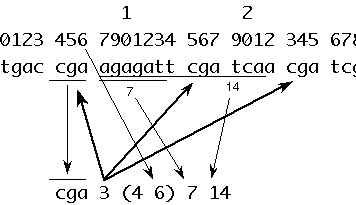
The long string output format is identical except that the string itself is not printed:
number_of_reps (first _location) first_distance sec_distance ...
This is easier to understand with an actual example. Recall that negative distance implies that the repeating substructure overlaps with its first instance.
A Test Example
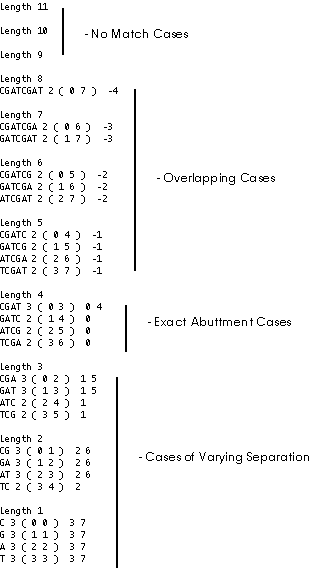
We will continue by factoring a sequence consisting of three repetitions of CTAG; CGATCGATCGAT is factored and those strings that occur more than once are printed.
testSequence statistics:
Stringlength: 12
Repeating Substrings: 62
Longest repeating substring(s):
Length 8: CGATCGAT 2 ( 0 7 ) -4
Longest repeating substring with no overlap:
Length 4: CGAT 3 ( 0 3 ) 0 4
Longest repeating substring with most repetitions, no overlap:
Length 4: CGAT 3 ( 0 3 ) 0 4
testSequence output:
With tested code it is now possible to answer the questions posed at the beginning.
Factoring L-19
L-19 was short enough to serve as a test and seemed to have relevance in the real world. Brookhaven and the National Institutes of Health maintain sequence data banks that one can access on the internet. I did so. The original annotated internet version of L-19 is included in Appendix A.
Input L-19
690 689 1
1 gggccgcagc catgagtatg ctcaggcttc agaagaggct cgcctctagt gtcctccgct
61 gtggcaagaa gaaggtctgg ttagacccca atgagaccaa tgaaatcgcc aatgccaact
121 cccgtcagca gatccggaag ctcatcaaag atgggctgat catccgcaag cctgtgacgg
181 tccattcccg ggctcgatgc cggaaaaaca ccttggcccg ccggaagggc aggcacatgg
241 gcataggtaa gcggaagggt acagccaatg cccgaatgcc agagaaggtc acatggatga
301 ggagaatgag gattttgcgc cggctgctca gaagataccg tgaatctaag aagatcgatc
361 gccacatgta tcacagcctg tacctgaagg tgaaggggaa tgtgttcaaa aacaagcgga
421 ttctcatgga acacatccac aagctgaagg cagacaaggc ccgcaagaag ctcctggctg
481 accaggctga ggcccgcagg tctaagacca aggaagcacg caagcgccgt gaagagcgcc
541 tccaggccaa gaaggaggag atcatcaaga ctttatccaa ggaggaagag accaagaaat
601 aaaacctccc actttgtctg tacatactgg cctctgtgat tacatagatc agccattaaa
661 ataaaacaag ccttaaaaaa aaaaaaaacc
Factored L-19 Statistics
Stringlength: 690
Repeating Substrings: 3545
Longest repeating substring(s):
Length 13 aaaaaaaaaaaaa 2 ( 674 686 ) -12
Longest repeating substring(s) with no overlap:
Length 9
gccaatgcc 2 ( 107 115 ) 147
aaaacaagc 2 ( 408 416 ) 245
aggcccgca 2 ( 456 464 ) 24
aaataaaac 2 ( 596 604 ) 53
Longest repeating substring with most repetitions, no overlap:
Length 7
caagaag 3 ( 64 70 ) 392 476
agaccaa 3 ( 93 99 ) 404 488
ggcccgc 3 ( 214 220 ) 236 269
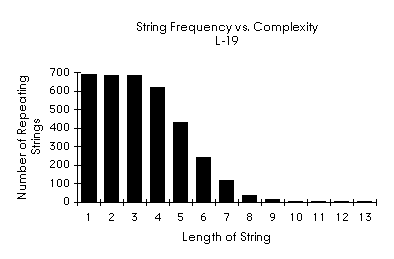
L-19 Observations
The longest repeating subunit with no overlap was 9 units long. This did not confirm the occurrence of repeating "micromachine" units I had hoped to find. This test did not take aliasing or wobble into account. Wobble allows for the substitution of various nucleic acids without changing the identity of the amino acid. Perhaps l-19 it is below the threshold of interesting substructures. An informative graph is :
A Random Example
It appears that the histogram above does not vary markedly from that which would be produced by an arbitrary substrings. A computer science colleague, Rod Bogart, suggested that the digits of p might be an interesting place to look for "longest repeating substrings". Instead of looking at English text or the digits of p I generated a random string whose length was the same as L-19. The results are interesting:
Randomly Generated String 690 Statistics
Stringlength: 690
Repeating Substrings: 3260
Longest repeating substring(s):
Length 9
gtgggggtg 2 ( 61 69 ) 211
tccgttgcc 2 ( 367 375 ) 152
ttcagactg 2 ( 471 479 ) 18
Longest repeating substring(s) with no overlap:
Length 9
gtgggggtg 2 ( 61 69 ) 211
tccgttgcc 2 ( 367 375 ) 152
ttcagactg 2 ( 471 479 ) 18
Longest repeating substring with most repetitions, no overlap:
Length 7
gggggtg 3 ( 63 69 ) 213
ctcggtc 3 ( 91 97 ) 11
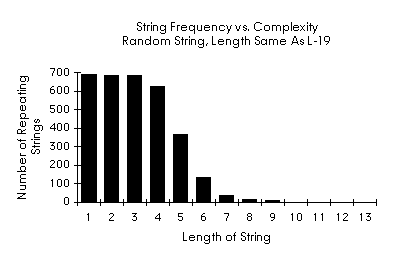
Random Observations
Surprisingly the randomly generated string shows organizational statistics that are strikingly similar in form to those of L-19. This tends, to a first approximation, to refute the presupposition that functions are organized by substructures of linear sequences.
Factoring Barley Hydrolase 612
The protein databank version of Barley Hydrolase are included in Appendix B. This is a synthetic example since it is known that this material consists of two distinct fragments, however it proves that multiple instances of complex structures can be found via the facString program. Amino acids were encoded into single letters by arbitrary substitution. Program facString does not depend on the encoding.
Encoding of Barley Hydrolase 612
ILE: I, GLY: G, VAL: V, CYS: C, TYR: T,
LEU: L, PRO: P, SER:S, ARG:A, ASP: B,
GLN: N, LYS:Y, MET:M, PHE:H, ALA:D,
THR: R, TRP:E, GLU:U, HIS:J
Input Barley Hydrolase 612
612 310 1
IGVCTGVIGAALPSASBVVNLTASYGIAGMAITHDBGNDLSDLAASGIGL
ILBIGABNLDAIDDSRSADDSEVNAAVAPTTPDVAIYTIDDGAUVNGGDR
NSILPDMAALADDLSDDGLGDIYVSRSIAHBUVDASHPPSDGVHYADTMR
BVDALLDSRGDPLLDAVTPTHDTABAPGSISLATDRHNPGRRVABNAAGL
RTRSLHBDMVBDVTDDLUYDGDPDVYVVVSUSGEPSDGGHDDSDGADART
ANGLIAJVGGGRPYYAUDLURTIHDMHAUANYRGBDRUASHGLHAPBYSP
DTAINH
IGVCTGVIGAALPSASBVVNLTASYGIAGMAITHDBGNDLSDLAASGIGL
ILBIGABNLDAIDDSRSADDSEVNAAVAPTTPDVAIYTIDDGAUVNGGDR
NSILPDMAALADDLSDDGLGDIYVSRSIAHBUVDASHPPSDGVHYADTMR
BVDALLDSRGDPLLDAVTPTHDTABAPGSISLATDRHNPGRRVABNAAGL
RTRSLHBDMVBDVTDDLUYDGDPDVYVVVSUSGEPSDGGHDDSDGADART
ANGLIAJVGGGRPYYAUDLURTIHDMHAUANYRGBDRUASHGLHAPBYSP
DTAINH
Barley Hydrolase 612 Statistics
Stringlength: 612
Repeating Substrings: 93,942
Longest repeating substring(s):
Length 306
2 ( 0 305 ) 0
Longest repeating substring(s) with no overlap:
Length 305
2 (0 304) 1
2 (1 305) 1
Longest repeating substring with most repetitions, no overlap:
Length 7
4 (38 41): 70 302 376
4 (138 141): 92 302 398
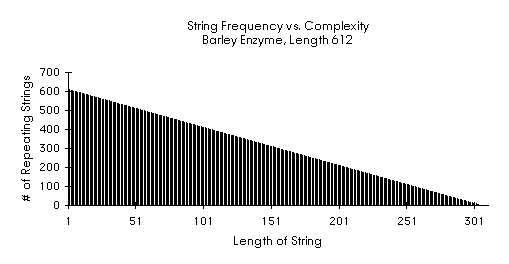
Barley Hydrolase Observations
Besides the obvious fact that facString finds the two identical substructures, the most conspicuous feature is that the shape of the histogram is linear, not sigmoidal as in the two previous cases. This linear shape comes from the fact that a repeating pattern of significant length has been detected.
Conclusions
L-19 appears for all intents and purposes to be no more sophisticated than its randomly generated counterpart. Repeated idiomatic expressions are common in computer programs and various other coded language. We see no definite evidence of repeated idiomatic expressions in L-19 when compared with a random control. This is surprising. The Barley Hydrolase example confirms that high level structures of significant complexity can be found using the technique.
Future Work
Further work includes running this on wider variety of more sophisticated sequences . Coding by amino acid rather than by nucleic acid reduces computer time requirements and accounts for aliasing. It would interesting to look at other plants, microorganisms, enzymes and viruses. It would also be informative to look for aliasing at higher levels of organization.
Looking at how other coding systems embed information might be useful in some kind of "Comparative Coding" or anatomy of coding schemes.
Since this all looks like codebreaking, it might be interesting to combine the expertise and resources of the NSA with those of the NIH and let the big guns have at it. ·
Acknowledgments
I got this idea when I was visiting Steve Mittelstaedt who was doing some pipetting one day in his office at UAMS. I asked him what he was doing and he said, "Sequencing" with a mystique reminiscent of the word "Plastics" whispered in The Graduate. I had come to his lab get some liquid nitrogen, and while waiting I noticed a chart of amino acids on his wall. "Sequencing" kept ringing in my head, it seemed so similar to "Text Processing" which I had done for several years in a computer science context. A few days later his mother, Bo Mittelstaedt, was kind enough to answer some basic questions and helped me articulate the notion of factoring sequences like symbolic expressions.
Appendix A: Internet Posting of L-19
LOCUS S56985 690 bp mRNA PRI 07-MAY-1993
DEFINITION ribosomal protein L19 [human, breast cancer cell line, MCF-7, mRNA,
690 nt].
ACCESSION S56985
KEYWORDS .
SOURCE human MCF-7 breast cancer cell line.
ORGANISM Homo sapiens
Unclassified.
REFERENCE 1 (bases 1 to 690)
AUTHORS Henry,J.L., Coggin,D.L. and King,C.R.
TITLE High-level expression of the ribosomal protein L19 in human breast
tumors that overexpress erbB-2
JOURNAL Cancer Res. 53, 1403-1408 (1993)
STANDARD full automatic
COMMENT GenBank staff at the National Library of Medicine created this
entry [NCBI gibbsq 127871] from the original journal article.
This sequence comes from Fig. 2.
NCBI gi: 298485
FEATURES Location/Qualifiers
source 1..690
/organism="Homo sapiens"
/note="human"
CDS 12..602
/note="Method: conceptual translation supplied by author.
This sequence comes from Fig. 2. NCBI gi: 298486"
/codon_start=1
/product="ribosomal protein L19"
/translation="MSMLRLQKRLASSVLRCGKKKVWLDPNETNEIANANSRQQIRKL
IKDGLIIRKPVTVHSRARCRKNTLARRKGRHMGIGKRKGTANARMPEKVTWMRRMRIL
RRLLRRYRESKKIDRHMYHSLYLKVKGNVFKNKRILMEHIHKLKADKARKKLLADQAE
ARRSKTKEARKRREERLQAKKEEIIKTLSKEEETKK"
BASE COUNT 216 a 175 c 184 g 115 t
ORIGIN
1 gggccgcagc catgagtatg ctcaggcttc agaagaggct cgcctctagt gtcctccgct
61 gtggcaagaa gaaggtctgg ttagacccca atgagaccaa tgaaatcgcc aatgccaact
121 cccgtcagca gatccggaag ctcatcaaag atgggctgat catccgcaag cctgtgacgg
181 tccattcccg ggctcgatgc cggaaaaaca ccttggcccg ccggaagggc aggcacatgg
241 gcataggtaa gcggaagggt acagccaatg cccgaatgcc agagaaggtc acatggatga
301 ggagaatgag gattttgcgc cggctgctca gaagataccg tgaatctaag aagatcgatc
361 gccacatgta tcacagcctg tacctgaagg tgaaggggaa tgtgttcaaa aacaagcgga
421 ttctcatgga acacatccac aagctgaagg cagacaaggc ccgcaagaag ctcctggctg
481 accaggctga ggcccgcagg tctaagacca aggaagcacg caagcgccgt gaagagcgcc
541 tccaggccaa gaaggaggag atcatcaaga ctttatccaa ggaggaagag accaagaaat
601 aaaacctccc actttgtctg tacatactgg cctctgtgat tacatagatc agccattaaa
661 ataaaacaag ccttaaaaaa aaaaaaaacc
//
Appendix B: Internet Posting of Barley Hydrolase
HEADER HYDROLASE 12-OCT-93 1GHS 1GHS 2
COMPND 1,3-BETA-GLUCANASE (E.C.3.2.1.39) 1GHS 3
COMPND 2 (1,3-BETA-D-GLUCAN ENDOHYDROLASE, ISOZYME II) 1GHS 4
SOURCE GERMINATED BARLEY GRAIN (HORDEUM VULGARE) 1GHS 5
AUTHOR T.P.J.GARRETT,J.N.VARGHESE 1GHS 6
REVDAT 1 01-NOV-94 1GHS 0 1GHS 7
JRNL AUTH J.N.VARGHESE,T.P.J.GARRETT,P.M.COLMAN,L.CHEN, 1GHS 8
JRNL AUTH 2 P.J.HOJ,G.B.FINCHER 1GHS 9
JRNL TITL THE THREE-DIMENSIONAL STRUCTURES OF TWO PLANT 1GHS 10
JRNL TITL 2 BETA-GLUCAN ENDOHYDROLASES WITH DISTINCT SUBSTRATE 1GHS 11
JRNL TITL 3 SPECIFICITIES 1GHS 12
JRNL REF PROC.NAT.ACAD.SCI.USA V. 91 2785 1994 1GHS 13
JRNL REFN ASTM PNASA6 US ISSN 0027-8424 0040 1GHS 14
REMARK 1 1GHS 15
REMARK 1 REFERENCE 1 1GHS 16
REMARK 1 AUTH L.CHEN,T.P.J.GARRETT,J.N.VARGHESE,G.B.FINCHER, 1GHS 17
REMARK 1 AUTH 2 P.B.HOJ 1GHS 18
REMARK 1 TITL CRYSTALLIZATION AND PRELIMINARY X-RAY ANALYSIS OF 1GHS 19
REMARK 1 TITL 2 (1,3)- AND (1,3;1,4)-BETA--D-GLUCANASES FROM 1GHS 20
REMARK 1 TITL 3 GERMINATING BARLEY 1GHS 21
REMARK 1 REF J.MOL.BIOL. V. 234 888 1993 1GHS 22
REMARK 1 REFN ASTM JMOBAK UK ISSN 0022-2836 0070 1GHS 23
REMARK 1 REFERENCE 2 1GHS 24
REMARK 1 AUTH L.CHEN,G.B.FINCHER,P.J.HOJ 1GHS 25
REMARK 1 TITL EVOLUTION OF POLYSACCHARIDE HYDROLASE SUBSTRATE 1GHS 26
REMARK 1 TITL 2 SPECIFICITY 1GHS 27
REMARK 1 REF J.BIOL.CHEM. V. 268 13318 1993 1GHS 28
REMARK 1 REFN ASTM JBCHA3 US ISSN 0021-9258 0071 1GHS 29
REMARK 2 1GHS 30
REMARK 2 RESOLUTION. 2.3 ANGSTROMS. 1GHS 31
REMARK 3 1GHS 32
REMARK 3 REFINEMENT. 1GHS 33
REMARK 3 PROGRAM X-PLOR 1GHS 34
REMARK 3 AUTHORS BRUNGER 1GHS 35
REMARK 3 R VALUE 0.179 1GHS 36
REMARK 3 RMSD BOND DISTANCES 0.013 ANGSTROMS 1GHS 37
REMARK 3 RMSD BOND ANGLE 1.68 DEGREES 1GHS 38
REMARK 3 1GHS 39
REMARK 3 NUMBER OF REFLECTIONS 22811 1GHS 40
REMARK 3 RESOLUTION RANGE 6.0 - 2.3 ANGSTROMS 1GHS 41
REMARK 3 DATA CUTOFF 4.0 SIGMA(F) 1GHS 42
REMARK 3 PERCENT COMPLETION 78.4 1GHS 43
REMARK 3 1GHS 44
REMARK 3 NUMBER OF PROTEIN ATOMS 4564 1GHS 45
REMARK 3 NUMBER OF SOLVENT ATOMS 59 1GHS 46
REMARK 4 1GHS 47
REMARK 4 THERE ARE TWO MOLECULES IN THE ASYMMETRIC UNIT. 1GHS 48
REMARK 4 THE TRANSFORMATION PRESENTED ON *MTRIX* RECORDS BELOW WILL 1GHS 49
REMARK 4 YIELD APPROXIMATE COORDINATES FOR CHAIN *A* WHEN APPLIED TO 1GHS 50
REMARK 4 CHAIN *B*. THE RMS DEVIATION IN CA POSITIONS IS 0.30 1GHS 51
REMARK 4 ANGSTROMS. 1GHS 52
REMARK 5 1GHS 53
REMARK 5 CIS PEPTIDES WERE OBSERVED IN THE ELECTRON DENSITY BETWEEN 1GHS 54
REMARK 5 RESIDUES PHE 274 AND ALA 275 FOR EACH MOLECULE. 1GHS 55
REMARK 6 1GHS 56
REMARK 6 THIS DEPOSITION CORRESPONDS TO THE INITIAL STRUCTURE 1GHS 57
REMARK 6 REPORT. DIFFRACTION DATA EXTEND TO ABOUT 1.8 A. 1GHS 58
REMARK 7 1GHS 59
REMARK 7 CROSS REFERENCE TO SEQUENCE DATABASE 1GHS 60
REMARK 7 SWISS-PROT ENTRY NAME PDB ENTRY CHAIN NAME 1GHS 61
REMARK 7 E13B_HORVU A 1GHS 62
REMARK 7 E13B_HORVU B 1GHS 63
SEQRES 1 A 306 ILE GLY VAL CYS TYR GLY VAL ILE GLY ASN ASN LEU PRO 1GHS 64
SEQRES 2 A 306 SER ARG SER ASP VAL VAL GLN LEU TYR ARG SER LYS GLY 1GHS 65
SEQRES 3 A 306 ILE ASN GLY MET ARG ILE TYR PHE ALA ASP GLY GLN ALA 1GHS 66
SEQRES 4 A 306 LEU SER ALA LEU ARG ASN SER GLY ILE GLY LEU ILE LEU 1GHS 67
SEQRES 5 A 306 ASP ILE GLY ASN ASP GLN LEU ALA ASN ILE ALA ALA SER 1GHS 68
SEQRES 6 A 306 THR SER ASN ALA ALA SER TRP VAL GLN ASN ASN VAL ARG 1GHS 69
SEQRES 7 A 306 PRO TYR TYR PRO ALA VAL ASN ILE LYS TYR ILE ALA ALA 1GHS 70
SEQRES 8 A 306 GLY ASN GLU VAL GLN GLY GLY ALA THR GLN SER ILE LEU 1GHS 71
SEQRES 9 A 306 PRO ALA MET ARG ASN LEU ASN ALA ALA LEU SER ALA ALA 1GHS 72
SEQRES 10 A 306 GLY LEU GLY ALA ILE LYS VAL SER THR SER ILE ARG PHE 1GHS 73
SEQRES 11 A 306 ASP GLU VAL ALA ASN SER PHE PRO PRO SER ALA GLY VAL 1GHS 74
SEQRES 12 A 306 PHE LYS ASN ALA TYR MET THR ASP VAL ALA ARG LEU LEU 1GHS 75
SEQRES 13 A 306 ALA SER THR GLY ALA PRO LEU LEU ALA ASN VAL TYR PRO 1GHS 76
SEQRES 14 A 306 TYR PHE ALA TYR ARG ASP ASN PRO GLY SER ILE SER LEU 1GHS 77
SEQRES 15 A 306 ASN TYR ALA THR PHE GLN PRO GLY THR THR VAL ARG ASP 1GHS 78
SEQRES 16 A 306 GLN ASN ASN GLY LEU THR TYR THR SER LEU PHE ASP ALA 1GHS 79
SEQRES 17 A 306 MET VAL ASP ALA VAL TYR ALA ALA LEU GLU LYS ALA GLY 1GHS 80
SEQRES 18 A 306 ALA PRO ALA VAL LYS VAL VAL VAL SER GLU SER GLY TRP 1GHS 81
SEQRES 19 A 306 PRO SER ALA GLY GLY PHE ALA ALA SER ALA GLY ASN ALA 1GHS 82
SEQRES 20 A 306 ARG THR TYR ASN GLN GLY LEU ILE ASN HIS VAL GLY GLY 1GHS 83
SEQRES 21 A 306 GLY THR PRO LYS LYS ARG GLU ALA LEU GLU THR TYR ILE 1GHS 84
SEQRES 22 A 306 PHE ALA MET PHE ASN GLU ASN GLN LYS THR GLY ASP ALA 1GHS 85
SEQRES 23 A 306 THR GLU ARG SER PHE GLY LEU PHE ASN PRO ASP LYS SER 1GHS 86
SEQRES 24 A 306 PRO ALA TYR ASN ILE GLN PHE 1GHS 87
SEQRES 1 B 306 ILE GLY VAL CYS TYR GLY VAL ILE GLY ASN ASN LEU PRO 1GHS 88
SEQRES 2 B 306 SER ARG SER ASP VAL VAL GLN LEU TYR ARG SER LYS GLY 1GHS 89
SEQRES 3 B 306 ILE ASN GLY MET ARG ILE TYR PHE ALA ASP GLY GLN ALA 1GHS 90
SEQRES 4 B 306 LEU SER ALA LEU ARG ASN SER GLY ILE GLY LEU ILE LEU 1GHS 91
SEQRES 5 B 306 ASP ILE GLY ASN ASP GLN LEU ALA ASN ILE ALA ALA SER 1GHS 92
SEQRES 6 B 306 THR SER ASN ALA ALA SER TRP VAL GLN ASN ASN VAL ARG 1GHS 93
SEQRES 7 B 306 PRO TYR TYR PRO ALA VAL ASN ILE LYS TYR ILE ALA ALA 1GHS 94
SEQRES 8 B 306 GLY ASN GLU VAL GLN GLY GLY ALA THR GLN SER ILE LEU 1GHS 95
SEQRES 9 B 306 PRO ALA MET ARG ASN LEU ASN ALA ALA LEU SER ALA ALA 1GHS 96
SEQRES 10 B 306 GLY LEU GLY ALA ILE LYS VAL SER THR SER ILE ARG PHE 1GHS 97
SEQRES 11 B 306 ASP GLU VAL ALA ASN SER PHE PRO PRO SER ALA GLY VAL 1GHS 98
SEQRES 12 B 306 PHE LYS ASN ALA TYR MET THR ASP VAL ALA ARG LEU LEU 1GHS 99
SEQRES 13 B 306 ALA SER THR GLY ALA PRO LEU LEU ALA ASN VAL TYR PRO 1GHS 100
SEQRES 14 B 306 TYR PHE ALA TYR ARG ASP ASN PRO GLY SER ILE SER LEU 1GHS 101
SEQRES 15 B 306 ASN TYR ALA THR PHE GLN PRO GLY THR THR VAL ARG ASP 1GHS 102
SEQRES 16 B 306 GLN ASN ASN GLY LEU THR TYR THR SER LEU PHE ASP ALA 1GHS 103
SEQRES 17 B 306 MET VAL ASP ALA VAL TYR ALA ALA LEU GLU LYS ALA GLY 1GHS 104
SEQRES 18 B 306 ALA PRO ALA VAL LYS VAL VAL VAL SER GLU SER GLY TRP 1GHS 105
SEQRES 19 B 306 PRO SER ALA GLY GLY PHE ALA ALA SER ALA GLY ASN ALA 1GHS 106
SEQRES 20 B 306 ARG THR TYR ASN GLN GLY LEU ILE ASN HIS VAL GLY GLY 1GHS 107
SEQRES 21 B 306 GLY THR PRO LYS LYS ARG GLU ALA LEU GLU THR TYR ILE 1GHS 108
SEQRES 22 B 306 PHE ALA MET PHE ASN GLU ASN GLN LYS THR GLY ASP ALA 1GHS 109
SEQRES 23 B 306 THR GLU ARG SER PHE GLY LEU PHE ASN PRO ASP LYS SER 1GHS 110
SEQRES 24 B 306 PRO ALA TYR ASN ILE GLN PHE 1GHS 111